Hvor mutasjoner feiler -del II
(fra 'Theistic Evolution - a Scientific, Philosophical and Theological critique, kap.7)
(fortsatt herfra)
III. Behovet for romlig informasjon
Etter at RNA og proteiner er dannet i en celle, må mange av dem transportert til ulike lokasjoner for å fungere skikkelig. I tillegg til deres protein-kodende regioner, har noen m-RNA sekvenser som kalles 'zip-koder' som spesifiserer adresser i cellen som de skal transporteres til. (35) Lik postnr i postvesener, er imidlertid zip-koden ubrukelig uten en pre-eksisterende adresse. Celler med kjerne inneholder mikroskopiske fibre, kalt "mikrotubuler". (Nano)molekylære maskiner ferdes langs mikrotubulene, og transporterer ulik last i gjennom hele cellen.
Om en benytter postnummer metaforen, så kan molekylære maskiner sammenlignes med varebiler og mikrotubulene med et motorvei-system. Men adressene for intracellulær transport, hvordan bestemmes de? I noen tilfeller kan adressesene spesifiseres som den romlige arrangering av motorveier. Ved å benytte postnummer metaforen igjen, kan noen varebiler bli ekspedert til en spesiell motorvei, og så fraktes til enden av veien, og losset. I andre tilfeller er destinasjonen kjent å være spesifisert ved molekyler innbygd i cellemembranen. Opprinnelig trodde vitenskapsfolk at proteiner kunne flyte fritt gjennom en membran, lik båter gjennom en kanal. Men nå er det imidlertid kjent at mange membran proteiner er arrangert i ikke-tilfeldige mønstre, som kan være ganske stabile. (36)
M.a.o. inneholder biologiske membraner romlig informasjon. Den informasjonen formidles delvis ved en sukker-kode og en bio-elektrisk kode.
A. Sukker-koden
Som vi så ovenfor overstiger den informasjonsbærende evne ved forgrenede glykaner mye den samme hos DNA og proteiner. Det finnes også lange ikke-forgrenede glykaner, kalt 'glyko-saminoglykaner' (GAGer) som primært inntreffer på overflaten til en celle. Selv om de ikke er forgrenede, kan GAG-er samles fra dusinvis av ulike underenheter, og sulfatgrupper kan tilknyttes dem i en vid variasjon av mønstre. Dette gjør GAGer til noen av de mest informasjonsrike molekyler i naturen. (37) Faktisk har informasjonen i forgrenede og ikke-forgrenede gykaner, blitt kalt "sukker-koden". (38)
Biolog R. Schnaar skrev i 1985 at det er "en kode på overflaten av hver celle, som spesifiserer dens funksjon og styrer samhandling med andre celler." Schnaar mistenkte at "sukker tjener som kodens alfabet". (39) Vi vet nå at han hadde rett. Overflaten av alle levende celler, studert til dags dato, er dekket med glykaner. Celle-overflate glykaner i tidlige embryoer som ormer, bananfluer, fisk, kylling og mus endrer seg på et høyst ordnet vis, og danner mønstre 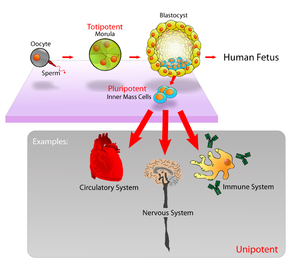 som er korresponderer med ulike stadier av embryo-utviklingen. De er involvert i celle-orientering, sammentrekning, bevegelse, svar på regulerende faktorer og celle-celle kommunikasjon.
som er korresponderer med ulike stadier av embryo-utviklingen. De er involvert i celle-orientering, sammentrekning, bevegelse, svar på regulerende faktorer og celle-celle kommunikasjon.
Sukkerkoden blir fortolket av proteiner som kalles 'lektiner', som gjenkjenner spesifikke tre-dimensjonale strukturer og glykaner, ved hjelp av 'karbohydrat-gjenkjennende-domener'. (40) Lektiner spesifiserer en bred variasjon av interaksjon mellom celler.
Bilde 8. Ulike celletyper -tidlige i embryo-utvikling
B. Den bio-elektriske koden
Cellens overflate-kode inkluderer mer enn sukker-koden. Det har lenge vært kjent at sannsynligvis alle levende celler genererer elektriske felt over sine membraner. (41) I dyre-celler flytter en natrium-kalium pumpe i membranen som frakter natriumceller ut av cella, mens den tar to kalium-ioner inn i den. Det øker det intracellulære trykket av kalium-ioner, som så flyter ut av celler gjennom ione-selektive kanaler i membranen. Den kombinerte virkningen fra natrium-kalium pumpen og kalium-'lekkasje' gjør at det indre av cellen får negativ elektrisk ladning i forhold til det ytre. Det resulterer i et endogent elektrisk felt.
Mønsteret av ione-kanaler i membranen bestemmer formene til de endogene elektriske feltene, som endres i løpet av embryo-utviklingen. Det endogene elektriske feltet i et froskeegg, driver en strøm av ioner gjennom egget. I det det befruktede egget deler seg, og cellene re-arrangerer seg for å danne et rumpetroll, endrer det elektriske feltet seg på organiserte måter. Det samme gjelder kylling-embryo. (42) I frosk, kylling og mus -embryoer, merkes fremtidige steder for lemmenes utvikling ved sterk, utgående strøm, før lemmene dukker opp der. (43)
Hvordan påvirker endogene elektriske felt embryo-utvikling? En måte er trolig ved å styre celle-bevegelser. Biologer har anvendt kunstige elektriske felt av fysiologisk styrke på embryoniske celler og observert at noen typer av celler flytter seg mot en den ene eller andre polen. Elektriske felt av fysiologisk styrke, kan også påvirke celledeling. (44) Sterke bevis for at endogene elektriske felt spiller betydelige roller i embryo-utviklingen kommer fra å kunstig ødelegge dem og så observere effektene. Å redusere en ione-strøm, assosiert med utvikling av av hale(stjert) utvikling i et kylling-embryo, resulterer i abnormaliteter. (45) Å manipulere ione-kanaler som genererer et elektrisk felt assosiert med øye-utvikling, i froske-embryoer, resulterer i deformerte eller manglende øyne, eller rumpetroll med øyne lokalisert på siden av halen. (46)
Så endogene elektriske felt er visselig involvert i embryo-utvikling. R. Shi og R. Borgens gikk gjennom det eksperimentelle bevis i 1995, og konkluderte med at at slike felt "kan forsyne et tre-dimensjonalt koordinat system" som hjelper å spesifisere form i embryoer. (47) M. Levin argumenterte at endogene elektriske felt kan fungere som "form-maler" og "det er sannsynlig at en full forståelse av [utviklingen] .. og dens informasjons-innehold, trenger å involvere å knekke den bio-elektriske koden." (48)
IV. Implikasjoner for neo-darwinismen
Så membran-mønstre bærer essensiell biologisk informasjon. Likevel kan ikke den informasjonen reduseres til sekvensinformasjonen i DNA. Selv om DNA-sekvenser fullstendig skulle spesifisere m-RNA og proteiner, så må den romlige distribusjonen av proteiner i celle-membranen spesifiseres uavhengig, lik adresser i post-metaforen. Så hva er det som spesifiserer membran-mønstre?
Bilde 9. Thylakoid-membran
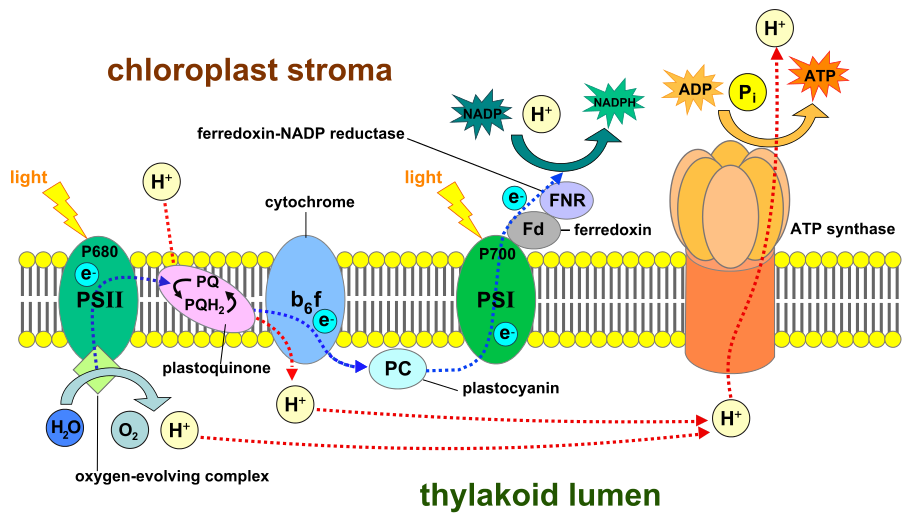 A. Membran-arv
A. Membran-arv
Noen membran mønstre har maler fra membranene de stammer fra i celledelingens løp. I slike tilfeller inkorporeres proteiner fra cellens indre, gjennom membranens vekst, bare om de matcher det eksisterende avtrykket (matrix -49). Biolog R. Poyton foreslo at biologiske membraner bærer 'romlig hukommelse', der enhetene er romlig lokaliserte multi-underenhets proteiner. I det en membran vokser i forberedelse for en deling, skilles proteinene fra hverandre inn i sine underavdelinger. Nylig syntetiserte enheter i cytoplasmaet forbinder seg med eldre underenheter for å danne hybride proteiner som er kjemisk identiske med de opprinnelige proteinene, og bevarer den opprinnelige romlige informasjonen. Poyton konkluderte: "Det å innse at genetisk hukommelse er én-dimensjonal langs ett DNA-molekyl, mens romlig hukommelse sannsynligvis er to-dimensjonal langs membranens overflate, og tre-dimensjonal i cellens indre, så er det sannsynlig at romlig hukommelse er mer komplisert og mangfoldig enn genetisk hukommelse." (50) Videre forskning har funnet bevis konsistent med Poytons hypotese. (51)
Celle-overflate maler har blitt grundig studert i encellede organismer (protoza). Noen protoza er dekket med cilia, små hår som slår rytmisk for fremdrift og ernæring. For mer enn 50 år siden oppdaget forskere at om de kunstig endret cilia-mønsteret, så kunne avkommet etter en celledeling arve det endrede mønsteret, selv om DNA var uendret. Siden da har cilia-protoza med kunstig endret mønster blitt stødig etablert i laboratorier gjennom tusener av generasjoner. (52)
St. Ng og J. Frankel studerte fenomenet og konkluderte i 1977: "Cellen som arkitekt benytter ikke bare genomisk informasjon for å produsere egnede bygnings-blokker, men arrangerer dem også i tillegg i følge tegninger som definert i den pre-eksisterende arkitekturen." (53) Slike maler er ikke begrenset til protoza. I 1990 studerte M. Locke mønstre i overhud til sommerfugl-celler, og konkluderte at deres arv "avhang av mer enn antall og type av molekyl. Mønster-duplisering involverer relativ posisjon og orientering," faktorer som ikke kan spesifiseres bare gjennom en base-sekvens. (54)
Celler med kjerne inneholder mange ulike typer av membraner, som Th. Cavalier-Smith kollektivt har betegnet "membranomen". Hver membran er karakterisert ikke bare ved sin kjemi, men også med sitt membran mønster. . Cavalier-Smith kalte membraner som oppsto ved vekst og deling av samme type for 'genetiske membraner'. Om noe av den genetiske membranen gikk tapt, kunne det sannsynligvis ikke bli gjendannet fra sine opprinnelige molekyler, selv om hele protein-kodende DNA-sekvenser var intakte, fordi den nødvendige romlige informasjonen var borte.
Bilde 10. Annen type celle-membran
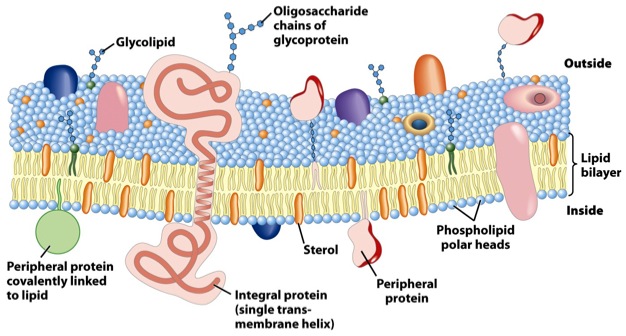
Så i følge Cavalier-Smith var ideen om at genomet inneholder all informasjon som behøves for å danne en organisme: "simpelt hen feilaktig. .. Dyrs utvikling danner en kompleks 3-dimensjonal multicellulær organisme, ikke ved å starte fra den lineære informasjon i DNA .. men alltid ved å starte med en allerede høyst kompleks tre-dimensjonal encellet organisme, det befruktede egg (55) Men membran-arv kan ikke være hele historien. I løpet av embryo-utvikling, dupliserer ikke bare membranen til celler de stammer fra; membran mønstre er meget ulike i nerveceller, muskelceller, blodceller, ben og lever -celler, bare for å nevne noen. Så membran mønstre må endres i løpet av embryo-utviklingen. Endringene er imidlertid ikke tilfeldige, de er høyst regulerte for å danne de integrerte organ-systemer til det voksne individet. Om membran mønstre ikke er spesifisert via membran-mønstre i celler de stammer fra, hvor er de så spesifisert?
B. En matematisk tilnærming
Det korte svaret er at vi ikke vet enda. Men teoretiske biologer har adressert problemet matematisk. I 1950 årene introduserte R. Rashevsky og R. Rosen en ny tilnærming for studiet av levende organismer, som de kalte "relasjonell biologi". I stedet for å gi ontologisk prioritet til molekyler, slik som DNA, gir relasjonell biologi forrang til relasjonene som konstituerer et organisert system. (56) For å gjøre tilnærmingen rigorøs, vendte Rashevsky og Rosen seg til "kategori-teorien" som var utviklet i 1940-årene. En generalisering av klassisk mengde-teori, kategori teori, som var blitt utviklet i 1940-årene -kan benyttes til modellere matematisk endringene av informasjon som inntreffer i levende organismer. Men det er ikke klart enda hvorvidt relasjonell biologi og kategori-teorien vil lykkes i å forklare endringer i løpet av embryo-utviklingen. Det som er klart, er at neo-darwinismen feiler i å forklare evolusjon.
C. Hvorfor neo-darwinismen feiler
I følge neo-darwinistisk teori forsyner DNA-mutasjoner råmaterialet til evolusjonen. Om det sentrale dogmet holdt mål, så kunne det være tilfelle. Men dogmet om at "DNA lager RNA som lager proteiner, som lager oss," feiler i hvert steg. Mange RNA blir modifisert til det nivået at de ikke lenger reflekterer base-sekvensen i DNA, som de ble transkribert fra. Og den endelige foldede formen til proteinet, som funksjonen avhenger av, blir i noen tilfeller ikke fullt 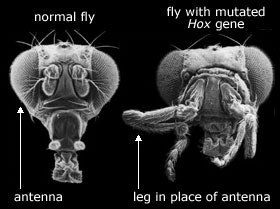 spesifisert av m-RNA som det blir oversatt fra. Videre blir de fleste proteiner ekstremt modifisert ved hjelp av glykosalisjon, tilføyelse av komplekse sukkerarter fra nettverk av enzymer. Så det endelige produkt av transkripsjon og oversettelse er vanligvis meget langt fra å bli spesifisert av DNA-sekvenser.
spesifisert av m-RNA som det blir oversatt fra. Videre blir de fleste proteiner ekstremt modifisert ved hjelp av glykosalisjon, tilføyelse av komplekse sukkerarter fra nettverk av enzymer. Så det endelige produkt av transkripsjon og oversettelse er vanligvis meget langt fra å bli spesifisert av DNA-sekvenser.
Ett enda mer alvorlig problem for sentraldogmet er at proteiner må bli skikkelig lokalisert i en celle for at den skal fungere. Men DNA spesifiserer ikke romlige koordinater. I stedet spesifiseres de ved arkitekturen til cellen og ved mønstre i membranen som går forut for transkripsjon og oversettelse. Så selv om DNA fullstendig spesifiserte proteinets endelige form, så måtte det enda være informasjon som var uavhengig av DNA.
Bilde 11. Mutasjon i hox-gen --neo-darwinismens bygningssten?
Det er sant at noen DNA-mutasjoner kan være tjenlige for en organisme i en spesiell omgivelse. F.eks. forgifter antibiotikaen streptomycin tuberkuløse bakterier ved å søke ut ribosomene deres og derved blokkere protein-syntese. For DNA-mutasjoner som delvis ødelegger ribosomene kan hindre antibiotikaen fra å gjenkjenne dem. Selv om de mutante bakteriene er ødelagt i nærvær av antibiotikaen, er de bedre stilt enn de ikke-ødelagte bakteriene.
Men å ødelegge et ribosom er meget forskjellig fra å produsere tjenlige nye variasjoner i embryo-utvikling, som kunne utgjøre råmaterialet for evolusjon. Biologer har søkt systematisk etter mutasjoner som påvirker utvikling av bananfluer, rundormer, sebrafisk og mus. Effektene av slike mutasjoner faller alltid i tre kategorier. Enten klarer embryoet å komme seg over endringene og utvikler seg normalt, eller så deformeres eller dør embryoet. Dette kan knapt kalles råmaterialet for evolusjon.
Referanser:
35. R.H. Singer, "RNA ZipCodes for CytoPlasmic Addresses," Current Biology 3 (1993): 719-721
36. D.M. Engelman, "Membranes are More Mosaic than Fluid," Nature 438 (2005): 578-580
37. J.E. Turmbull, "Heparan Sulfate Glycomics: Towards Systems Biology Strategics," BioChemical Society translationg 38 (2010): 1356-60
38. H-J. Gabius, "Biological Information Transfer Beyond the Genetic Code: The Sugar Code", Naturwissenschaften (2000): 108-121
39. R.L: Schnaar, "The Membrane is the Message," The Sciences, May-June 1985); 34-40
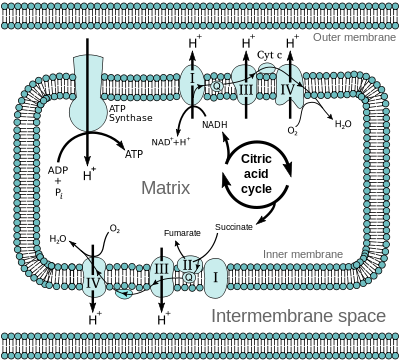
Bilde 12. Elektrontransport i mitokondriene
40. K. Drickamer, "Two Distinct Classes of Carbohydrate-Recognition Somains in Animal Lectins," Journal of Biological Chemistry 263 (1988): 9557-9560
41. H.S. Burr et al, "Evidence of the Existence of an ElectroDynamic Field in Living Organisations," Proceedings of the National Academy of Sciences USA 25 (1939): 284-288
42. R. Nutticelli, " Endogenous Electric Fields in Embryos during Development Regeneration and Wound Healing," Radiation Protection Dosimetry 106 (2003):375-383
43. A.M. Altizer et al., " Endogenous Electric Current Is Associated With Normal Developmen of the Vertebrate Limb," Dev. Dynamics 223 (2001):391-401
44. C.D. Mc Caig et al, "Electric Dimensions in Cell Science," Journal of Cell Science 122 (2009): 4267-4276
45. K.B. Hotary et al, "Evidence for a Role for Endogenous Electric Fields in Chick Embryo Development," Development 114 (1992): 985-996
46. V.P. Pai et al, "Transmembrane Voltage Portential Controls Embryonic Eye Patterning in Xenopus Laevis," Dev. 139 (2012): 313-323
47. Riyi Shi og R.B. Borgens, "Three-Dimensional Gradients of Voltage during Developmen of the Nervous System as Invisible Coordinates for the Establishment of Embryonic Pattern," Dev. Dynamics 202 (1995): 101-114
48. M. Levin, "Morphogenesic Fields in Embryogenesis, Regeneration and Cancer Non-Local Control of Complex Patterning," BioSystems 109 (2012): 243-261
50. R. O. Poyton, "Memory and Membranes: The Expression of Genetic that Is Specified Independently of DNA" BIO-Complexity 2014, no.2 (2014): 1-28
51. J. Wells, "Membrane Patterns Carry Ontogenous Information that is Specified Independently of DNA," BIO-Complexity 2014 no.2 (2014). 1-28
52. G.M. Grimes et al, Cellular Aspects of Pattern Formation: The Problem of Assembly (Basel: Karger, 1991)
53. St. F. Ng et al, "180grades Rotation of Cliary Rows and Its Morphogenetic Implications in Tetrahymena pyriformis,2 Proceedings of the National Academy of Sciences USA 74 (1977): 1115-1118
54. M: Locke, "Is There Somatic Inheritance of Intracellular Patterns?" Journal of Cell Science 96 (1990): 563-567
55. Th. Cavalier-Smith, "The Memranome and Membrane Heredity in Development and Evolution," In Organelles, Genomes and Eucaryotic Phylogeny, ed. R.P. Hirt and D.S. Horner (Boca Raton, FL: CRC Press, 2004), 335-351
56. R. Rosen, Life Itselv (NY: Columbia University Press, 1991)
Ansvarlig for oversettelse og bilder ved Asbjørn E. Lund
{kind=link}